A especiação
A especiação é sobre como as espécies se formam. É uma parte importante da biologia evolutiva. Darwin pensava que a maioria das espécies surgia diretamente de espécies pré-existentes. Isto é chamado de anagenese: espécie por mudança, ou "evoluçã…
A especiação é sobre como as espécies se formam. É uma parte importante da biologia evolutiva.
Darwin pensava que a maioria das espécies surgia diretamente de espécies pré-existentes. Isto é chamado de anagenese: espécie por mudança, ou "evolução filótica". Durante grande parte do século XX pensávamos que a maioria das espécies surgia por divisões de espécies anteriores: cladogênese. A visão geral era de que a maioria das divisões de espécies é causada ou ajudada em seu caminho por mecanismos de isolamento.
Sem dúvida, a separação física das espécies que uma vez viveram juntas é um fator principal. É ilustrada por tantos exemplos, alguns dos quais são discutidos abaixo.
No entanto, o trabalho dos últimos 20 anos tem mostrado algumas outras causas. A análise da seqüência de DNA dos seres vivos tem mostrado que muitas vezes existe alguma hibridação entre espécies relacionadas. Isso significa que os genes foram transferidos por esses cruzamentos. Por sua vez, isso significa que o isolamento reprodutivo não é a única definição de uma espécie, e a especiação nem sempre precisa de alopatia (espécie a ser separada reprodutivamente). As seções abaixo ilustram a idéia de que a separação física foi de primordial importância na formação de novas espécies.
Galeria de imagens
10 Imagens









Mecanismos de isolamento
Os mecanismos de isolamento são coisas que impedem a reprodução bem sucedida entre grupos de uma espécie. O isolamento reprodutivo das populações é estabelecido. Isto é particularmente importante para o conceito de espécies biológicas, pois as espécies são definidas pelo isolamento reprodutivo.
Os mecanismos de isolamento podem ser divididos em dois grupos, antes e depois da fertilização.
Antes da fertilização
Fatores que impedem o acasalamento dos indivíduos.
- Isolamento geográfico: As espécies ocorrem em áreas diferentes, e muitas vezes são separadas por barreiras.
- Isolamento temporal: Os indivíduos não acasalam porque estão ativos em momentos diferentes. Podem ser momentos diferentes do dia ou estações diferentes. Os períodos de acasalamento das espécies podem não coincidir. Os indivíduos não se encontram durante seus períodos de acasalamento, ou não se encontram de forma alguma.
- Isolamento ecológico: Os indivíduos só acasalam em seu habitat preferido. Eles não encontram indivíduos de outras espécies com preferências ecológicas diferentes.
- Isolamento comportamental: Indivíduos de diferentes espécies podem se encontrar, mas não se reconhece nenhum sinal sexual que possa ser dado. Um indivíduo escolhe um membro de sua própria espécie na maioria dos casos.
- Isolamento mecânico: Pode ser tentada a cópula, mas a transferência de esperma não ocorre. Os indivíduos podem ser incompatíveis devido ao tamanho ou à morfologia.
- Incompatibilidade do jogo: A transferência de esperma ocorre, mas o óvulo não é fertilizado.
Após a fertilização
Fatores que impedem o sucesso do acasalamento, tais como incompatibilidade genética, inviabilidade híbrida ou esterilidade.
- Mortalidade zigótica: O ovo é fertilizado, mas o zigoto não se desenvolve.
- Inviabilidade híbrida: Forma embrionária híbrida, mas não é viável.
- Esterilidade híbrida: O híbrido é viável, mas o adulto resultante é estéril.
- Avaria híbrida: Os híbridos de primeira geração (F1) são viáveis e férteis, mas outras gerações híbridas (F2 e backcrosses) são inviáveis ou estéreis.

Isolamento geográfico
Pensa-se que esta seja a causa mais comum da especiação. A primeira pessoa a pensar nisso foi Moritz Wagner, um explorador alemão e historiador natural.
O início da carreira de Wagner foi como geógrafo, e ele publicou uma série de livros geográficos sobre o Norte da África, o Oriente Médio e a América Tropical. Ele também foi um naturalista e colecionador aguçado, e é por este trabalho que ele é mais conhecido entre os biólogos. Ernst Mayr, o evolucionista e historiador da biologia, deu um relato do significado de Wagner. p562-565
Durante seus três anos na Argélia, Wagner (entre outras atividades) estudou os escaravelhos sem vôo Pimelia e Melasoma. Cada gênero é dividido em várias espécies, cada uma delas confinada a um trecho da costa norte entre os rios que descem das montanhas do Atlas até o Mediterrâneo. Assim que se atravessa um rio, aparece uma espécie diferente, mas intimamente relacionada.
"... uma espécie incipiente só [surgirá] quando alguns poucos indivíduos transgridirem as fronteiras limitantes de seu alcance... a formação de uma nova raça nunca será bem sucedida... sem uma longa separação contínua entre os colonos e os outros membros de sua espécie".
Esta foi uma descrição antecipada de um processo de um tipo de especiação geográfica. Em 1942 foi reintroduzido por Mayr, e a importância da especiação geográfica tornou-se uma das idéias centrais da síntese evolucionária.
Outro termo para a especiação geográfica é especiação alopátrica. Allopatry significa "terra diferente".
Ilhas vagas
As ilhas vulcânicas são formadas sem vida, e toda a vida tem que chegar levada pelo vento ou pela água. Sabemos desde as ilhas havaianas e desde as ilhas Galápagos que todas as formas de vida mudam quando chegam às ilhas a partir do continente.
Em cerca de 6.500 km2 (17.000 km2), as ilhas havaianas possuem a mais diversa coleção de moscas drosófilas do mundo, vivendo de florestas tropicais a prados de montanha. São conhecidas cerca de 800 espécies de drosófilos havaianos.
Estudos mostram um claro "fluxo" de espécies de ilhas mais antigas para ilhas mais novas. Há também casos de colonização de volta às ilhas mais antigas, e de saltos de ilhas, mas estes são muito menos freqüentes.
Por datação de potássio/argônio radioativo, as ilhas atuais datam de 0,4 milhões de anos atrás (mya) (Mauna Kea) a 10mya (Necker). O membro mais antigo do arquipélago havaiano ainda acima do mar é o Atol de Kure, que pode ser datado até 30 mya.
O próprio arquipélago, produzido pela placa do Pacífico movendo-se sobre um ponto quente, existe há muito mais tempo, pelo menos até o Cretáceo. As ilhas havaianas mais as antigas ilhas que agora estão sob o mar compõem a cadeia de montanhas marinhas Havaianas-Emperadoras; e muitas das montanhas submarinas são manobras.
Todas as espécies nativas drosófilas em Hawaiʻi descendem aparentemente de uma única espécie ancestral que colonizou as ilhas, cerca de 20 milhões de anos atrás. A radiação adaptativa subsequente foi estimulada pela falta de competição e por uma grande variedade de nichos vagos. Embora fosse possível para uma única fêmea grávida colonizar uma ilha, é mais provável que tenha sido um grupo da mesma espécie.
Há outros animais e plantas no arquipélago havaiano que sofreram radiações adaptativas similares, ainda que menos espetaculares.
Espécies de anéis
Em biologia, uma espécie de anel é uma série conectada de populações vizinhas, cada uma das quais pode se cruzar com populações vizinhas. As duas extremidades da cadeia se sobrepõem.
As duas populações finais da série estão muito distantes da reprodução cruzada. Tais populações "finais", não cruzadas, embora ligadas geneticamente, podem coexistir na mesma região, fechando assim um "anel".
As espécies de anéis fornecem importantes evidências de evolução: elas ilustram o que acontece ao longo do tempo à medida que as populações divergem geneticamente. Richard Dawkins observou que as espécies de anéis "só nos mostram na dimensão espacial algo que deve sempre acontecer na dimensão temporal".
Entretanto, é difícil encontrar um exemplo simples e direto.
Gaivotas de Larus
Um exemplo clássico de espécies de anéis é o anel de espécies circumpolares das gaivotas Larus. A variedade destas gaivotas forma um anel ao redor do Pólo Norte, que normalmente não é sobrevoado por gaivotas individuais.
Em termos gerais, as gaivotas podem cruzar em certa medida com seus vizinhos, exceto nas duas extremidades do anel.
As gaivotas de fundo preto e as gaivotas de arenque são suficientemente diferentes que normalmente não se hibridizam; assim, o grupo de gaivotas forma um continuum, exceto onde as duas linhagens se encontram na Europa.
Embora a realidade seja muito mais complicada, ela mostra bem a idéia geral.
Salamandras Ensatina
A salamandra Ensatina é uma espécie de anel nas montanhas ao redor do Vale Central da Califórnia. O complexo forma uma ferradura em forma de ferradura ao redor das montanhas. Embora a reprodução cruzada possa ocorrer entre cada uma das 19 populações ao redor da ferradura, a subespécie Ensatina eschscholtzii na extremidade oeste da ferradura não pode cruzar com a Ensatina klauberi na extremidade leste. É uma ilustração de "quase todas as etapas de um processo de especiação" (Dobzhansky). Richard Highton argumentou que Ensatina é um caso de múltiplas espécies e não um continuum de uma espécie.
A Toutinegra Esverdeada
O Greenish Warbler (Phylloscopus trochiloides) tem uma série de subespécies, das quais P. t . viridianus é a mais familiar na Europa. É uma espécie de anel com populações divergindo para o leste e oeste do planalto tibetano, encontrando-se posteriormente no lado norte. Suas relações são bastante confusas.




Especiação simpátrica
A especiação simpátrica refere-se à formação de duas ou mais espécies descendentes de uma única espécie ancestral, todas ocupando a mesma localização geográfica. Pensa-se agora que isto seja muito comum.
Na especiação simpática, as espécies divergem ao habitarem o mesmo lugar. Exemplos frequentemente citados de especiação simpátrica são encontrados em insetos que se tornam dependentes de diferentes plantas hospedeiras na mesma área.
A existência da especiação simpátrica como um mecanismo de especiação foi muito contestada. As pessoas argumentaram que as evidências da especiação simpátrica são, de fato, exemplos de especiação microgeográfica. Em geral, esta não é agora a explicação preferida. Ao contrário, é o resultado da hibridização entre espécies estreitamente relacionadas, seguida da seleção natural trabalhando na progênie de tais cruzamentos. Um exemplo amplamente aceito de especiação simpátrica é o dos ciclideos do Lago Nabugabo na África Oriental, que se pensa ser devido à seleção sexual.
Especiação via poliploidização
A poliploidia tem causado muitos eventos de especiação rápida porque os descendentes de, por exemplo, acasalamentos tetraplóides x diplóides muitas vezes resultam em progênie estéril triplóide.
Entretanto, nem todas as poliplóides estão reprodutivamente isoladas de suas plantas parentais, e o fluxo gênico ainda pode ocorrer, por exemplo, através de acasalamentos híbridos triplóides x diplóides que produzem tetraplóides.
Muitas das espécies vegetais existentes e a maioria das espécies animais aparentemente sofreram polioidização em sua história evolutiva. A reprodução de espécies poliplóides de sucesso é às vezes assexuada, por partenogênese. Por razões desconhecidas, muitos organismos assexuais são poliploides.
Mosca Hawthorn
Um exemplo de evolução no trabalho é o caso da mosca espinheiro, Rhagoletis pomonella, que parece estar sofrendo uma especiação simpática.
Diferentes populações de moscas de espinheiro se alimentam de diferentes frutas. Uma população distinta surgiu na América do Norte no século 19, algum tempo depois que as maçãs, uma espécie não nativa, foram introduzidas. Esta população de maçãs alimenta-se normalmente apenas de maçãs e não das frutas historicamente preferidas dos espinheiros. A população atual de espinheiros não se alimenta normalmente de maçãs.
Algumas evidências sugerem que a especiação simpátrica está ocorrendo. As moscas Hawthorn amadurecem mais tarde na estação e levam mais tempo para amadurecer do que as moscas da maçã; e há poucas evidências de reprodução cruzada (pesquisadores documentaram uma taxa de hibridação de 4-6%).
O surgimento da nova mosca espinheiro é um exemplo de evolução em andamento.
Hibridação
Raramente uma nova espécie se forma quando membros individuais de diferentes espécies acasalam. Normalmente, os produtos de tais acasalamentos híbridos são inférteis (não férteis), ou relativamente, e por isso são eliminados pela seleção natural.
Um exemplo de uma nova e bem sucedida espécie híbrida foi encontrado. A nova espécie é um híbrido dos pardais italianos e espanhóis, e os fatos básicos foram verificados por análise de seqüência a partir do DNA em seu sangue. A ave vive na Itália, em uma região onde vivem as duas espécies-mãe. Ela não se reproduz com o pardal espanhol, apesar de viver com eles lado a lado.
Especiação artificial
Novas espécies foram criadas pela criação de animais domésticos, mas as datas e métodos iniciais não são claros. Por exemplo, ovinos domésticos foram criados por hibridização, e não produzem mais descendência viável com Ovis orientalis, uma espécie da qual são descendentes.
O gado doméstico, por outro lado, pode ser considerado a mesma espécie que várias variedades de boi selvagem, gaur, iaque, etc., uma vez que produzem prontamente descendência fértil com eles.
Espécies de laboratório
As criações mais bem documentadas de novas espécies no laboratório foram realizadas no final dos anos 80. William Rice e G.W. Salt criaram moscas da fruta, Drosophila melanogaster, usando um labirinto com três diferentes opções de habitat, como luz/escuro e úmido/seco.
Cada geração foi colocada no labirinto, e os grupos de moscas que saíram de duas das oito saídas foram separados para procriar um com o outro em seus respectivos grupos. Após trinta e cinco gerações, os dois grupos e seus descendentes foram isolados reprodutivamente por causa de suas fortes preferências de habitat: acasalaram apenas dentro das áreas que preferiam, e assim não acasalaram com moscas que preferiam as outras áreas. A história de tais tentativas é descrita em Rice and Hostert (1993).
Diane Dodd também foi capaz de mostrar como o isolamento reprodutivo pode se desenvolver a partir das preferências de acasalamento em Drosophila pseudoobscura após apenas oito gerações usando diferentes tipos de alimentos, amido e maltose.
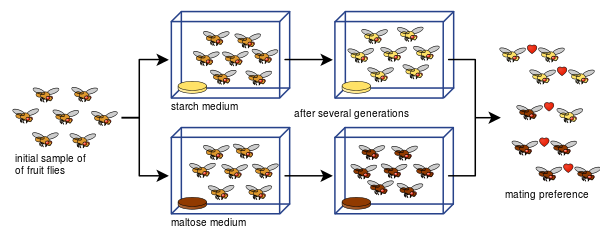
A experiência de Dodd tem sido fácil de replicar para muitos outros, inclusive com outros tipos de moscas da fruta e alimentos.
Reforço (efeito Wallace)
Reforço é o processo pelo qual a seleção natural aumenta o isolamento reprodutivo.
Pode ocorrer depois que duas populações da mesma espécie são separadas e depois voltam a entrar em contato. Se seu isolamento reprodutivo foi completo, então eles já terão se desenvolvido em duas espécies separadas e incompatíveis.
Se seu isolamento reprodutivo for incompleto, então o acasalamento posterior entre as populações produzirá híbridos, que podem ou não ser férteis. Se os híbridos forem inférteis, ou férteis, mas menos aptos que seus ancestrais, então não haverá mais isolamento reprodutivo e a especiação ocorreu essencialmente (por exemplo, como nos cavalos e burros).
O raciocínio por trás disso é que se os pais da descendência híbrida tiverem, cada um, características naturalmente selecionadas para seus próprios ambientes diferentes, a descendência híbrida terá características de ambos, e não se encaixaria em nenhum dos nichos, assim como em nenhum dos pais. A baixa aptidão dos híbridos faria com que a seleção favorecesse o acasalamento sortido, o que reduziria a hibridação.
Isto é às vezes chamado de efeito Wallace depois que o biólogo evolucionista Alfred Russel Wallace sugeriu no final do século XIX que poderia ser um fator importante na especiação.
Se os descendentes híbridos forem mais aptos que seus ancestrais, então as populações se fundirão novamente na mesma espécie dentro da área em que estão em contato.
O reforço é necessário para a separação quando existe uma "zona híbrida" entre duas formas de uma espécie. As zonas híbridas são regiões onde populações divergentes se encontram e intercruzam. As crias híbridas são muito comuns nestas regiões, que geralmente são criadas por espécies divergentes que entram em contato secundário.
Sem reforço, as duas populações ou espécies teriam cruzamentos incontroláveis. O reforço pode ser induzido em experimentos de seleção artificial, como descrito acima.
Perguntas e respostas
P: O que é a especiação?
R: A especiação é o processo de como as espécies se formam. É uma parte importante da biologia evolutiva.
P: O que Darwin pensou sobre a formação das espécies?
R: Darwin pensava que a maioria das espécies vinha diretamente de espécies pré-existentes, o que ele chamou de anagenesis ou "evolução filótica".
P: Como era geralmente vista a especiação no século XX?
R: No século XX, os cientistas pensavam que a maioria das espécies se formava quando as espécies anteriores se dividiam, o que era conhecido como cladogênese. Também se acreditava que essa divisão era causada ou ajudada por mecanismos de isolamento, tais como a separação física.
P: Como as pesquisas recentes mudaram nosso entendimento sobre a especiação?
R: Pesquisas recentes mostraram que a hibridação entre espécies relacionadas pode ocorrer e que os genes podem ser transferidos através desses cruzamentos. Isso significa que o isolamento reprodutivo nem sempre é necessário para definir uma espécie e a alopatia (separação física) pode não ser necessária para que a especiação ocorra.
P: O que a separação física tem a ver com a formação de novas espécies?
R: Acredita-se que a separação física entre espécies uma vez coexistentes seja um fator principal na formação de novas espécies, ilustrado por muitos exemplos discutidos na literatura científica.
Artigos relacionados
Autor
AlegsaOnline.com A especiação Leandro Alegsa
URL: https://pt.alegsaonline.com/art/92570
Fontes
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833